
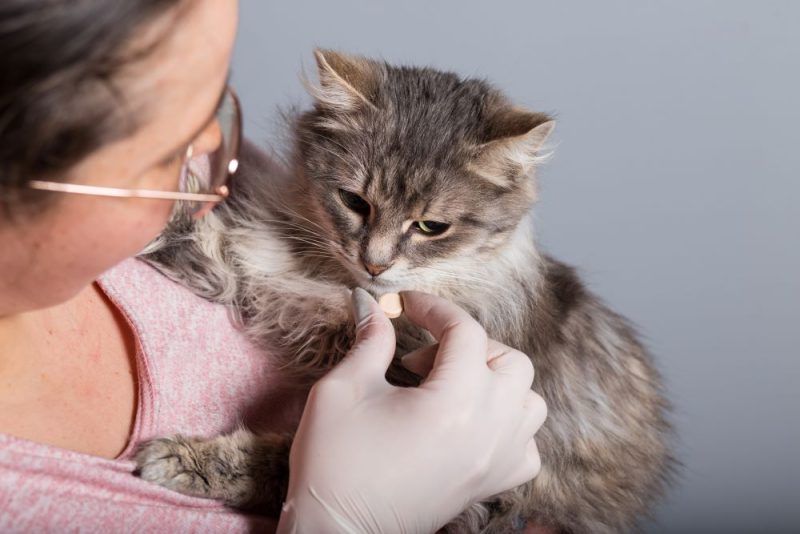
Tabletka na kleszcze, którą kot zjada ze smakiem? To możliwe!
Zabezpieczenie kota przed pasożytami zewnętrznymi jest bardzo ważne, zwłaszcza jeśli nasz pupil wychodzi na spacery lub ma dostęp do ogródka. Jednym z najskuteczniejszych i najwygodniejszych preparatów profilaktycznych są tabletki dla kota na pchły i kleszcze. Często jednak u właścicieli pojawiają…

Czy pies może jeść Rutinoscorbin z witaminą C? Szkodliwy czy nie?
Kiedy nasz futrzany przyjaciel błagalnie patrzy na nas, gdy otwieramy kolejną saszetkę witamin, nasuwa się pytanie – czy pies może jeść Rutinoscorbin? W końcu jako opiekunowie, zależy nam na ich zdrowiu tak samo, jak na własnym. Czy jednak to, co…

Czy pies może jeść mąkę pszenną lub razową? Sprawdzamy!
Czy zastanawialiście się kiedyś, przechodząc obok kuchennych półek, czy pies może jeść mąkę, którą wykorzystujecie do wypieków? Czy produkt tak powszechny jak mąka pszenna lub razowa może stać się częścią psiego menu? W tym artykule rozwiejemy wszystkie wątpliwości i pokażemy,…

Czy pies może jeść rzeżuchę? Szkodliwa czy nie?
Czy zastanawialiście się kiedyś, podczas przygotowywania wiosennego śniadania, czy wasz czworonożny przyjaciel może podzielić z wami radość z zielonych dodatków na talerzu? „Czy pies może jeść rzeżuchę?” – to pytanie, które może pojawić się w głowie każdego odpowiedzialnego właściciela psa.…

Czy pies może jeść żelatynę? Szkodliwa czy nie?
Żelatyna to składnik, który często spotykamy w naszej kuchni, ale czy pies może jeść żelatynę? Ta substancja, popularna w wielu deserach, może wydawać się nieszkodliwa, jednak jej spożycie przez naszych czworonożnych przyjaciół budzi wiele kontrowersji. Czy jest szkodliwa dla zdrowia…
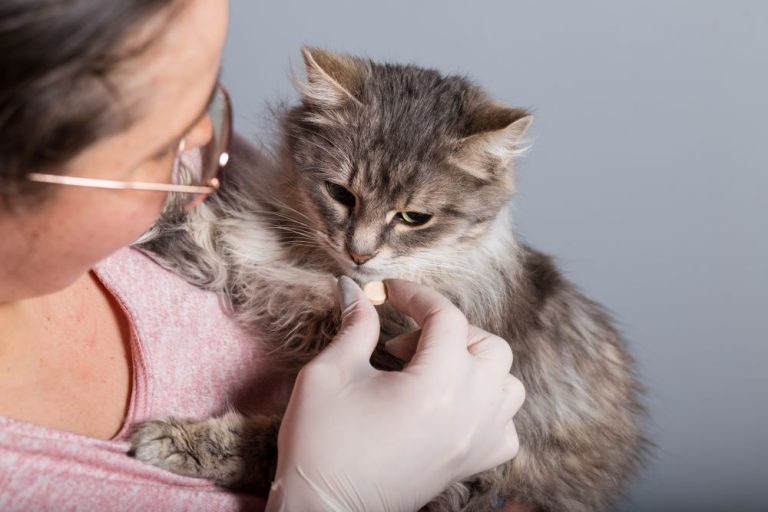
Tabletka na kleszcze, którą kot zjada ze smakiem? To możliwe!
Zabezpieczenie kota przed pasożytami zewnętrznymi jest bardzo ważne, zwłaszcza jeśli nasz pupil wychodzi na spacery lub ma dostęp do ogródka.…
Czy pies może jeść Rutinoscorbin z witaminą C? Szkodliwy czy nie?
Kiedy nasz futrzany przyjaciel błagalnie patrzy na nas, gdy otwieramy kolejną saszetkę witamin, nasuwa się pytanie – czy pies może…
Czy pies może jeść mąkę pszenną lub razową? Sprawdzamy!
Czy zastanawialiście się kiedyś, przechodząc obok kuchennych półek, czy pies może jeść mąkę, którą wykorzystujecie do wypieków? Czy produkt tak…
Czy pies może jeść rzeżuchę? Szkodliwa czy nie?
Czy zastanawialiście się kiedyś, podczas przygotowywania wiosennego śniadania, czy wasz czworonożny przyjaciel może podzielić z wami radość z zielonych dodatków…
Czy pies może jeść żelatynę? Szkodliwa czy nie?
Żelatyna to składnik, który często spotykamy w naszej kuchni, ale czy pies może jeść żelatynę? Ta substancja, popularna w wielu…

Psy z długimi uszami – TOP 10 psów z dużymi uszami
Generuję tekst

Najsłodsze psy świata – TOP 10 słodkich piesków
Kto może oprzeć się rozkosznemu wzrokowi i merdającemu ogonkowi? Zanurz się w świecie czterech łap, gdzie uśmiech nigdy nie znika…
Wysunięte prącie u psa – co oznacza i jak leczyć?
Jeśli zauważyłeś, że Twojemu czworonożnemu przyjacielowi niespodziewanie „psu wyszedł cały siusiak”, może to wywołać u Ciebie zaniepokojenie lub zdumienie. Zrozumienie…

Czemu pies liże mi nogę lub między nimi? Właściwości psiej śliny
Kiedy Twój czworonożny przyjaciel zaczyna czułościowo liżeć Ci nogę, lub nawet gdy pies mnie liże między nogami, możesz zastanawiać się…

Czy pies może jeść orzechy arachidowe? Sprawdzamy!
Czy zastanawiałeś się kiedyś, przechodząc obok półki ze smakołykami dla swojego czworonożnego przyjaciela, czy pies może jeść orzechy arachidowe? Te…

Czy pies może jeść bitą śmietanę? Sprawdzamy!
Czy zastanawialiście się kiedyś, podczas popołudniowej kawy z kremowym dodatkiem, czy psia miska mogłaby zostać wzbogacona o ten puszysty przysmak?…

Czy pies może jeść śnieg i lód? Sprawdzamy!
Podczas zimowych spacerów nasze czworonożne pociechy często zachowują się jak dzieci, bawiąc się w śniegu. Ale czy zastanawialiście się kiedyś,…